 Yihan Xiao1,2†
Yihan Xiao1,2† Xiuji Cui
Xiuji Cui- 1School of Basic Medicine and Life Science, Hainan Medical University, Haikou, Hainan, China
- 2Department of Clinical Laboratory, The Second Affiliated Hospital of Hainan Medical University, Haikou, Hainan, China
- 3Department of Laboratory Medicine, Hainan Affiliated Hospital of Hainan Medical University, Haikou, Hainan, China
- 4Department of Infectious Disease, The First Affiliated Hospital of Hainan Medical University, Haikou, Hainan, China
- 5Key Laboratory of Tropical Translational Medicine of Ministry of Education, Hainan Medical University, Haikou, Hainan, China
- 6Hainan Medical University-The University of Hong Kong Joint Laboratory of Tropical Infectious Diseases, Hainan Medical University, Haikou, Hainan, China
Spliced hepatitis B virus (HBV) RNA has been reported to affect the viral life cycle and progression of liver disease. As much as 30% of HBV RNA are spliced in infected cells, and some can be reverse transcribed to spliced DNA. To date, only a small number of spliced DNA have been identified and the majority require further investigation. This study aimed to identify the types of spliced DNA from the sera of a chronic hepatitis B patient. HBV DNA was isolated from the serum, and both full-length and spliced DNA were amplified by PCR. The amplified PCR products were then subcloned and sequenced using a Sanger sequence analysis. As a result, 19 types of spliced DNA were identified, 11 of which were assumed to be putative novel spliced DNA. These spliced DNA were doubly spliced with varied splice donor sites and splice acceptor sites. In addition to the frequently detected HBV SP1, SP2, and SP4, we also identified 11 putative novel spliced DNA in the serum. These findings indicate the complexity of HBV RNA splicing during viral life cycle.
Introduction
It has been estimated that approximately 7.18% of the population of China are Hepatitis B virus (HBV) carriers, and are thus at risk of developing liver cirrhosis and hepatocellular carcinoma (HCC) (Chen et al., 2006; Chen et al., 2013; Wang et al., 2014; Rapti and Hadziyannis, 2015). Thus, the HBV infection represents a public health problem in China.
HBV is a hepatotropic DNA virus with a partially double-stranded genome, also known as relaxed circular (rc) DNA (Delius et al., 1983). Upon entry into hepatocytes, the rcDNA is transported to the nucleus to form covalently closed circular (ccc) DNA by completing the positive strand and ligating gaps in the positive and negative DNA strands, respectively (Tuttleman et al., 1986; Hu and Seeger, 2015). The cccDNA functions as a template for all HBV viral RNA, including pregenomic (pg) RNA, precore RNA, preS1/preS2/S mRNA and X mRNA. Among these RNAs, pgRNA encodes the core protein and HBV reverse transcriptase, and also serves as the template for HBV DNA synthesis (Summers and Mason, 1982).
Unlike human immunodeficiency virus-1 and human papilloma virus, splicing of HBV RNA does not appear to be required in the HBV life cycle (Su et al., 1989a; Karn and Stoltzfus, 2012; Graham and Faizo, 2017; Kremsdorf et al., 2021). However, spliced HBV RNA has frequently been detected in chronic hepatitis B patient’s liver tissue (Su et al., 1989b; Bayliss et al., 2013; Chen et al., 2015). Some spliced RNAs have been shown to be further reverse transcribed into HBV DNA and subsequently secreted as defective HBV particles (Chen et al., 2015; Kremsdorf et al., 2021). This phenomenon has also been observed in an in vitro cell culture system (Abraham et al., 2008; Chen et al., 2015). Moreover, several studies have reported that the expression of isoforms and the amount of HBV splice variants, including spliced RNA and DNA vary in patients, transfected cell types, and HBV genotypes (Abraham et al., 2008; Chen et al., 2015; Kremsdorf et al., 2021; Lim et al., 2021). Moreover, these variants have been reported to modulate HBV replication and hepatocyte apoptosis and are associated with a poor response to interferon treatment and development of HCC (Chen et al., 2015; Betz-Stablein et al., 2016). However, the precise clinical and virological relevance of HBV splice variants remains poorly understood.
Currently, there are at least 20 pgRNA-derived HBV splice variants, including spliced RNA and DNA that have been detected in HBV infected liver tissue, serum, and transfected cells (Kremsdorf et al., 2021). However, the high proportion of HBV RNA splice variants among total HBV RNA production coupled with abundant HBV daily production (approximately 1011 copies/day) suggests currently reported HBV splice variants may only account for a small portion of the total variants (Lau and Wright, 1993; Nowak et al., 1996).
Thus, the aim of the study is to investigate the molecular characteristics of HBV spliced DNA in clinical serum sample. Here, we report 11 putative novel HBV spliced DNAs from the sera of a chronic hepatitis B patient.
Materials and methods
Isolation and amplification of serum HBV DNA
HBV DNA was isolated from 200 µL of serum of a 36-year-old male chronic hepatitis B patient, who had not been received antiviral treatment yet and was negative for hepatitis C virus, using a QIAamp MinElute Virus Spin Kit (Qiagen, German) in accordance with the manufacturer’s instructions. The full-length HBV genome was amplified using forward primer P1and reverse primer P2 with KOD DNA polymerase (TOYOBO, Japan), which possesses proof-reading activity, to minimize nucleotide mismatch. The sampling procedures were approved by the Ethics Committee of the Hainan Medical University, Haikou, Hainan province, China.
Clonal analysis
PCR products including approximately 3.2kb-long DNA (full-length HBV DNA) band and near 2kb-long DNA (short fragment HBV DNA) were recovered from a 1% agarose gel, ligated with the pLB-vector (TIANGEN, China), and subsequently transformed into DH5α Escherichia coli according to standard protocol. Positive colonies detected with conventional colony PCR with P1/P2 primers were further cultured in Luria Broth medium containing ampicillin (100 mg/mL) at 37°C for 16 h. Plasmids were isolated by TIANprep mini plasmid kit (TIANGEN, China) and subjected to HBV genome sequencing using ABI 3730xl DNA analyzer (Sangon Biotech, China). Primers for the sequencing of full-length (primer T7, P4, P5, P6, and P7) and short fragment (primer T7, RT-R, and pLB-R) HBV DNA are listed in Table 1 and the primer binding site is described in Figure 1 (Gunther et al., 1998).
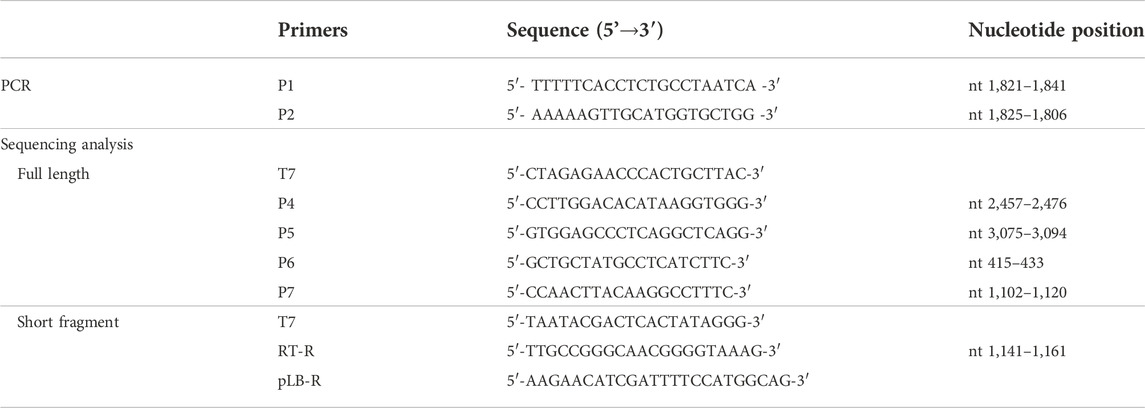
TABLE 1. List of primer pairs.
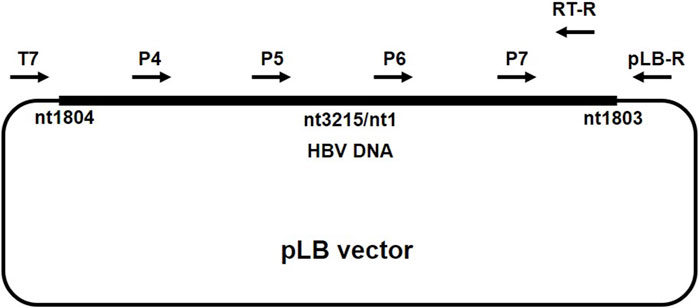
FIGURE 1. Primer binding site. The position of primers used in the sequencing of full-length HBV DNA and short fragment HBV DNA.
Sequence analysis
The alignment of the obtained HBV sequences was performed using the ClustalW method and the HBV genotype was determined with a neighbor-joining phylogenetic tree using Molecular Evolutionary Genetics Analysis (MEGA) software, version X. Reference sequences obtained from GenBank were used to compare the HBV full-genome sequences from the present study. Accession numbers: AB697490, GQ358158, DQ069801, HM011493, EU410080, EU670263, GU721029, AP011106, AP011109, AB540583, AB554019, and AB554025.
Results
Amplification of full-length and short fragment of HBV genome
To study the types of secreted HBV spliced DNA, HBV genome isolated from the serum was amplified by PCR using primer pairs P1/P2. In addition to full length HBV genome, a large amount of approximately 2 kb-long short fragments were detected in the sera of a chronic hepatitis B patient (Figure 2, lane 2). The short fragment was assumed to be HBV spliced DNA due to the similar molecular size of a previously reported HBV splice variant (Kremsdorf et al., 2021).
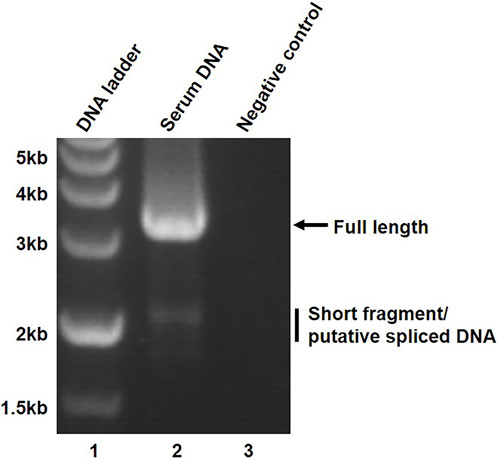
FIGURE 2. Amplification of full-length and short fragment of HBV genome. The amplified serum HBV DNA was resolved on a 1% agarose gel and the PCR products were visualized on UV light.
Identification of HBV splice variants
To confirm the assumption and identify the position of the genetic deletion in the short fragment, 50 colonies of subcloned short fragments were subjected to a sequencing analysis.
The obtained sequences were aligned with the full-length HBV DNA sequence (accession number: MA12345), which was determined to be HBV genotype C5 by a neighbor-joining phylogenetic tree analysis from the same PCR reaction. As a result, we detected 19 types of spliced DNA, including 8 previously reported types and 11 that have not yet been reported (Figure 3). Among these types, SP1 (42%, 21/50) was the most abundant HBV spliced DNA, followed by SP4 (10%, 5/50), SP2 (8%, 4/50) and SP5 (4%, 2/50) (Figure 3). There were 11 putative spliced DNA (pSD) that have not been previously reported, which were denoted pSD1, pSD2 …, pSD11 (Figure 3).
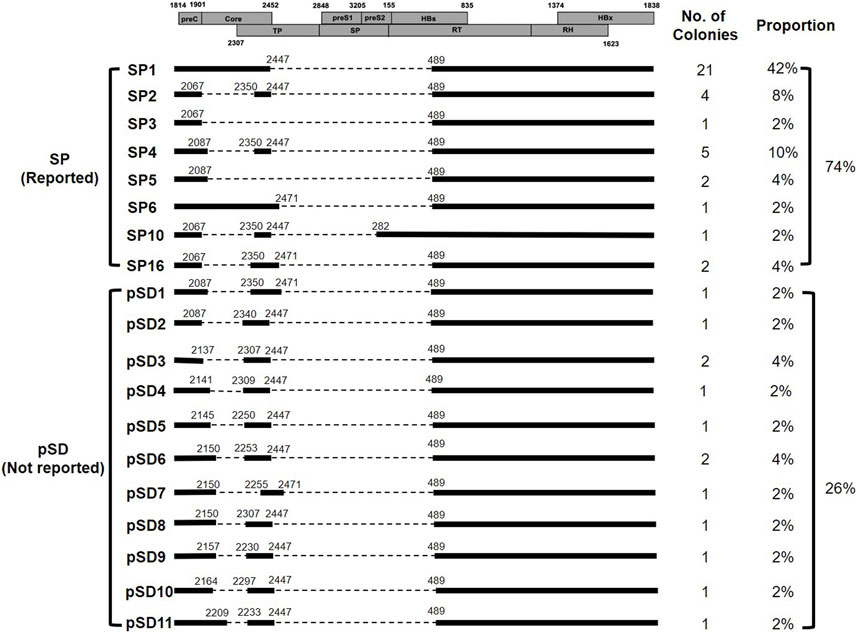
FIGURE 3. HBV spliced DNAs identified in the serum. Schematic diagram of the ORF of HBV genome (boxes), types of spliced DNAs including previously reported spliced DNA, denoted SP and putative novel spliced DNA, denoted pSD, and the proportion of the individual spliced DNA.
All spliced DNAs, except SP10, had a common splice donor at nucleotide (nt) 489 and splice acceptor at nt 2,447 or nt 2,471, however, various non-canonical splicing sites were also detected in pSDs (Table 2).
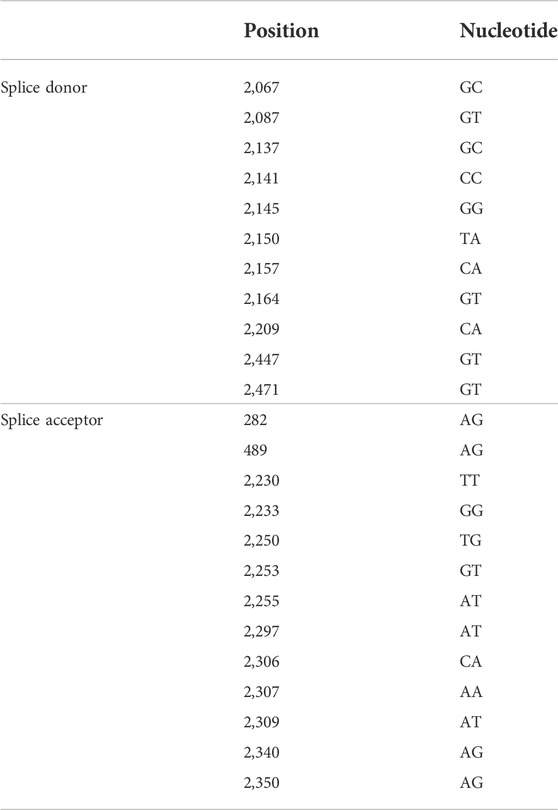
TABLE 2. Genetic variability of splice donor and splice acceptor.
Discussion
Splicing of HBV RNA, especially pgRNA splicing, frequently occurs during HBV replication, and the spliced pgRNA can be packaged into capsid to be transcribed into HBV DNA (Kremsdorf et al., 2021). According to previous studies, the proportion of intracellular HBV spliced RNA could comprise up to 30% of the total HBV RNA in cells transfected with HBV expressing plasmid construct in vitro, whereas the spliced DNA only accounted for approximately 1% of the total secreted HBV DNA in the sera (Chen et al., 2015; Lim et al., 2021). This discrepancy may be related to the lower efficiency of the assembly or secretion of viral particles with defective HBV DNA. Due to the small amount of secretion, spliced DNA is not easily detected during whole HBV genome amplification using conventional PCR method. However, during our routine study of HBV genome amplification, we detected a large amount of approximately 2kb-long PCR products that were assumed to be spliced DNA from the serum of a chronic hepatitis B patient. This finding provides us with an opportunity to study the molecular characteristics of spliced DNA.
In the present study, 19 types of HBV splice variants were detected, including 8 that have previously been reported and 11 putative novel spliced DNA (Su et al., 1989b; Chen et al., 1989; Abraham et al., 2008; Ma et al., 2009; Lim et al., 2021). Consistent with previous studies, SP1 was found to be the most abundant spliced DNA, accounting for 42% of the total variants (Liaw, 2013; Chen et al., 2015; Lim et al., 2021). In addition, we also detected SP2 (8%), SP3 (2%), SP4 (10%), SP5 (4%), SP6 (2%), SP10 (2%) and SP16 (4%) in various proportions.
All pSDs had common sties of splice donor and splice acceptor in the second region of deletion (from nt 2,447/2,471 to nt 489), however, they had varied splicing sites that were closely located to the canonical splice donor and splice acceptor sites at the first region of deletion (from nt 2,067 to nt 2,350). Moreover, unlike the previously reported consensus sequences of the splice sites (GT/GC for splice donor and AG for splice acceptor), TA/AT/TG was frequently detected in the splice donor site and the splice acceptor site of pSD (Kremsdorf et al., 2021). These findings suggest that nucleotide sequences near the canonical splice site may also contribute to HBV pgRNA splicing.
It has been previously reported that both HBV RNA and DNA variants are associated with the progression of liver disease and have a biological impact on the viral life cycle. SP1 is the most commonly studied HBV splice variant that encodes the HBV splice-generated protein (HBSP). The HBSP has been shown to inhibit HBV nucleocapsid formation, suppress host apoptosis, and impair the response to interferon treatment (Chen et al., 2015; Wang et al., 2015; Wu et al., 2018). In addition to SP1, the SP7, SP14, and SP19 have been reported to translate truncated HBV protein, which was assumed to affect the HBV viral life cycle (Huang et al., 2000; Park et al., 2008; Chen et al., 2010; Lin et al., 2012; Lin and Chen, 2017). In the present study, SP1 was the most abundantly detected HBV variant, whereas SP7, SP14, and SP19 were not detected.
Similar to SP2, SP4, SP10 and SP16, all pSDs were doubly spliced DNA, however, there are limited studies on the biological properties of doubly spliced variants. It has been reported that SP10 can suppress HBV transcription (Tsai et al., 2015). Considering that pSDs exhibited a similar pattern of splicing to that of SP10, they may have an impact on the HBV viral life cycle and pathogenesis, however, the precise mechanism requires further study.
Taken together, we identified multiple pSDs with varied splice donor and splice acceptor sites from the serum of chronic hepatitis B patient. These findings imply the complexity of HBV RNA splicing in the viral life cycle. A limitation of this study is the lack of liver tissue from the same patient for a comparison of the splice variants profile between the serum and liver tissue.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding authors.
Ethics statement
The studies involving human participants were reviewed and approved by Ethics Committee of the Hainan Medical University, Haikou, Hainan province, China. The patients/participants provided their written informed consent to participate in this study.
Author contributions
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
Funding
This work was supported by Emerging Tropical Infectious Diseases Science Innovation Group of Hainan Medical University, Hainan Provincial Natural Science Foundation of China (Grant No. 820RC640 for XC; No. 819MS122 for LS), the National Natural Science Foundation of China (Grant No. 81660335, 82060365 for XC).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
ccc, covalently closed circular; HBV, hepatitis B virus; HBSP, HBV splice-generated protein; HCC, hepatocellular carcinoma; nt, nucleotide; pSD, putative spliced DNA; rc, relaxed circular; SP, spliced protein.
References
Abraham, T. M., Lewellyn, E. B., Haines, K. M., and Loeb, D. D. (2008). Characterization of the contribution of spliced RNAs of Hepatitis B virus to DNA synthesis in transfected cultures of Huh7 and HepG2 cells. Virology 379, 30–37. doi:10.1016/j.virol.2008.06.021
Bayliss, J., Lim, L., Thompson, A. J., Desmond, P., Angus, P., Locarnini, S., et al. (2013). Hepatitis B virus splicing is enhanced prior to development of hepatocellular carcinoma. J. Hepatol. 59, 1022–1028. doi:10.1016/j.jhep.2013.06.018
Betz-Stablein, B. D., Topfer, A., Littlejohn, M., Yuen, L., Colledge, D., Sozzi, V., et al. (2016). Single-molecule sequencing reveals complex genome variation of hepatitis B virus during 15 Years of chronic infection following liver transplantation. J. Virol. 90, 7171–7183. doi:10.1128/jvi.00243-16
Chen, C. J., Yang, H. I., Su, J., Jen, C. L., You, S. L., Lu, S. N., et al. (2006). Risk of hepatocellular carcinoma across a biological gradient of serum Hepatitis B virus DNA level. JAMA 295, 65–73. doi:10.1001/jama.295.1.65
Chen, J., Wu, M., Wang, F., Zhang, W., Wang, W., Zhang, X., et al. (2015). Hepatitis B virus spliced variants are associated with an impaired response to interferon therapy. Sci. Rep. 5, 16459. doi:10.1038/srep16459
Chen, P. J., Chen, C. R., Sung, J. L., and Chen, D. S. (1989). Identification of a doubly spliced viral transcript joining the separated domains for putative protease and reverse transcriptase of Hepatitis B virus. J. Virol. 63, 4165–4171. doi:10.1128/jvi.63.10.4165-4171.1989
Chen, W., Zheng, R., Zhang, S., Zhao, P., Li, G., Wu, L., et al. (2013). The incidences and mortalities of major cancers in China, 2009. Chin. J. Cancer 32, 106–112. doi:10.5732/cjc.013.10018
Chen, W. N., Chen, J. Y., Lin, W. S., Lin, J. Y., and Lin, X. (2010). Hepatitis B doubly spliced protein, generated by a 2.2 kb doubly spliced Hepatitis B virus RNA, is a pleiotropic activator protein mediating its effects via activator protein-1- and CCAAT/enhancer-binding protein-binding sites. J. Gen. Virol. 91, 2592–2600. doi:10.1099/vir.0.022517-0
Delius, H., Gough, N. M., Cameron, C. H., and Murray, K. (1983). Structure of the Hepatitis B virus genome. J. Virol. 47, 337–343. doi:10.1128/jvi.47.2.337-343.1983
Graham, S. V., and Faizo, A. A. A. (2017). Control of human papillomavirus gene expression by alternative splicing. Virus Res. 231, 83–95. doi:10.1016/j.virusres.2016.11.016
Gunther, S., Sommer, G., Von Breunig, F., Iwanska, A., Kalinina, T., Sterneck, M., et al. (1998). Amplification of full-length Hepatitis B virus genomes from samples from patients with low levels of viremia: Frequency and functional consequences of PCR-introduced mutations. J. Clin. Microbiol. 36, 531–538. doi:10.1128/jcm.36.2.531-538.1998
Hu, J., and Seeger, C. (2015). Hepadnavirus genome replication and persistence. Cold Spring Harb. Perspect. Med. 5, a021386. doi:10.1101/cshperspect.a021386
Huang, H. L., Jeng, K. S., Hu, C. P., Tsai, C. H., Lo, S. J., and Chang, C. (2000). Identification and characterization of a structural protein of Hepatitis B virus: A polymerase and surface fusion protein encoded by a spliced RNA. Virology 275, 398–410. doi:10.1006/viro.2000.0478
Karn, J., and Stoltzfus, C. M. (2012). Transcriptional and posttranscriptional regulation of HIV-1 gene expression. Cold Spring Harb. Perspect. Med. 2, a006916. doi:10.1101/cshperspect.a006916
Kremsdorf, D., Lekbaby, B., Bablon, P., Sotty, J., Augustin, J., Schnuriger, A., et al. (2021). Alternative splicing of viral transcripts: The dark side of HBV. Gut 70, 2373–2382. doi:10.1136/gutjnl-2021-324554
Lau, J. Y., and Wright, T. L. (1993). Molecular virology and pathogenesis of Hepatitis B. Lancet 342, 1335–1340. doi:10.1016/0140-6736(93)92249-s
Liaw, Y. F. (2013). Reversal of cirrhosis: An achievable goal of Hepatitis B antiviral therapy. J. Hepatol. 59, 880–881. doi:10.1016/j.jhep.2013.05.007
Lim, C. S., Sozzi, V., Littlejohn, M., Yuen, L. K. W., Warner, N., Betz-Stablein, B., et al. (2021). Quantitative analysis of the splice variants expressed by the major Hepatitis B virus genotypes. Microb. Genom 7, mgen000492. doi:10.1099/mgen.0.000492
Lin, C. M., Wang, G. M., Jow, G. M., and Chen, B. F. (2012). Functional analysis of Hepatitis B virus pre-s deletion variants associated with hepatocellular carcinoma. J. Biomed. Sci. 19, 17. doi:10.1186/1423-0127-19-17
Lin, Y. M., and Chen, B. F. (2017). A putative Hepatitis B virus splice variant associated with chronic hepatitis and liver cirrhosis. Virology 510, 224–233. doi:10.1016/j.virol.2017.07.025
Ma, Z. M., Lin, X., Wang, Y. X., Tian, X. C., Xie, Y. H., and Wen, Y. M. (2009). A double-spliced defective Hepatitis B virus genome derived from hepatocellular carcinoma tissue enhanced replication of full-length virus. J. Med. Virol. 81, 230–237. doi:10.1002/jmv.21393
Nowak, M. A., Bonhoeffer, S., Hill, A. M., Boehme, R., Thomas, H. C., and McDade, H. (1996). Viral dynamics in Hepatitis B virus infection. Proc. Natl. Acad. Sci. U. S. A. 93, 4398–4402. doi:10.1073/pnas.93.9.4398
Park, G. S., Kim, H. Y., Shin, H. S., Park, S., Shin, H. J., and Kim, K. (2008). Modulation of Hepatitis B virus replication by expression of polymerase-surface fusion protein through splicing: Implications for viral persistence. Virus Res. 136, 166–174. doi:10.1016/j.virusres.2008.05.005
Rapti, I., and Hadziyannis, S. (2015). Risk for hepatocellular carcinoma in the course of chronic Hepatitis B virus infection and the protective effect of therapy with nucleos(t)ide analogues. World J. Hepatol. 7, 1064–1073. doi:10.4254/wjh.v7.i8.1064
Su, T. S., Lai, C. J., Huang, J. L., Lin, L. H., Yauk, Y. K., Chang, C. M., et al. (1989a). Hepatitis B virus transcript produced by RNA splicing. J. Virol. 63, 4011–4018. doi:10.1128/jvi.63.9.4011-4018.1989
Su, T. S., Lui, W. Y., Lin, L. H., Han, S. H., and P'eng, F. K. (1989b). Analysis of Hepatitis B virus transcripts in infected human livers. Hepatology 9, 180–185. doi:10.1002/hep.1840090203
Summers, J., and Mason, W. S. (1982). Replication of the genome of a Hepatitis B-like virus by reverse transcription of an RNA intermediate. Cell 29, 403–415. doi:10.1016/0092-8674(82)90157-x
Tsai, K. N., Chong, C. L., Chou, Y. C., Huang, C. C., Wang, Y. L., Wang, S. W., et al. (2015). Doubly spliced RNA of hepatitis B virus suppresses viral transcription via TATA-binding protein and induces stress granule assembly. J. Virol. 89, 11406–11419. doi:10.1128/jvi.00949-15
Tuttleman, J. S., Pourcel, C., and Summers, J. (1986). Formation of the pool of covalently closed circular viral DNA in hepadnavirus-infected cells. Cell 47, 451–460. doi:10.1016/0092-8674(86)90602-1
Wang, F. S., Fan, J. G., Zhang, Z., Gao, B., and Wang, H. Y. (2014). The global burden of liver disease: The major impact of China. Hepatology 60, 2099–2108. doi:10.1002/hep.27406
Wang, Y. L., Liou, G. G., Lin, C. H., Chen, M. L., Kuo, T. M., Tsai, K. N., et al. (2015). The inhibitory effect of the Hepatitis B virus singly-spliced RNA-encoded p21.5 protein on HBV nucleocapsid formation. PLoS One 10, e0119625. doi:10.1371/journal.pone.0119625
Keywords: hepatitis B virus, chronic hepatitis B, spliced DNA, splice protein, hepatocellular carcinoma
Citation: Xiao Y, Cui Z, Zhou X, Li Z, Li L, Wu L, Sun L and Cui X (2023) Molecular characterization of multiple putative novel hepatitis B virus spliced DNA from a chronic hepatitis B patient. Acta Virol. 67:11663. doi: 10.3389/av.2023.11663
Received: 20 May 2022; Accepted: 17 February 2023;
Published: 04 July 2023.
Edited by:
Katarina Polcicova, Slovak Academy of Sciences, SlovakiaCopyright © 2023 Xiao, Cui, Zhou, Li, Li, Wu, Sun and Cui. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiuji Cui, Y3VpeGoyNkBoYWlubWMuZWR1LmNu; Long Sun, aHkwMjAzMDY4QGhhaW5tYy5lZHUuY24=
†These authors have contributed equally to this work