 Ian O. Bledsoe1*
Ian O. Bledsoe1* Melanie A. Morrison2
Melanie A. Morrison2- 1Department of Neurology, Weill Institute for Neurosciences, University of California, San Francisco, San Francisco, CA, United States
- 2Department of Radiology and Biomedical Imaging, University of California, San Francisco, San Francisco, CA, United States
Much remains to be learned about the mechanism of benefit of deep brain stimulation in movement disorders in general and dystonia specifically. A full accounting of the pathophysiology of dystonia additionally remains unclear. Given its ability to evaluate whole-brain network changes, functional neuroimaging is an important tool to advance understanding of the effects of deep brain stimulation, which in turn could offer insight into the pathophysiology of dystonia and suggest novel deep brain stimulation targets for the disorder. This review surveys the published literature of functional neuroimaging studies evaluating deep brain stimulation effects in dystonia, including PET, SPECT, and functional MRI studies. To date, study cohorts have been relatively small, though several general patterns emerge when studies are viewed collectively, including reduced functional activation patterns with stimulation turned on during motor tasks, particularly in frontal cortical regions. During rest with stimulation on, several studies showed areas of relatively decreased perfusion only in those participants who experienced clinical benefit from deep brain stimulation. Future research may benefit from larger cohorts with more homogeneous forms of dystonia, potentially enabled by multi-center initiatives. Additional benefits may result from more detailed longitudinal assessments and greater use of functional MRI, with study designs that take into account the technical limitations of this modality in the context of movement disorders and deep brain stimulation.
Introduction
The mechanism of benefit of deep brain stimulation (DBS) for dystonia remains unclear. Recent advances, including implanted devices that have the capability of sensing neural signals in addition to stimulating [1], have the potential to significantly broaden our understanding of DBS effects on abnormal motor networks in dystonia. While such devices offer superb temporal resolution and allow evaluations in the real world rather than only laboratory conditions, one limitation is their restricted geographic reach, sampling regions only directly adjacent to recording electrodes. With increasing recognition that dystonia is a network disorder [2], the ability to probe the distant neurophysiologic effects of DBS is of paramount importance for a full accounting of its mechanism. Functional neuroimaging can help fill this critical niche given its ability to assay large geographic regions simultaneously. Such an increased geographical reach might serve multiple purposes. First, it could help clarify the full extent of network changes from DBS, which would improve both the understanding of the mechanism of DBS effects and the underlying pathophysiology of dystonia. Second, it could potentially identify novel DBS targets in dystonia by evaluating the degree to which stimulation-induced changes in connected regions of a functional network are correlated with the greatest DBS benefit.
Despite such potential benefits to a better understanding of DBS mechanisms and dystonia pathophysiology, the published literature to date on functional imaging of DBS effects in dystonia is relatively sparse. Early studies utilized molecular brain imaging techniques including positron emission tomography (PET) and single photon emission computed tomography (SPECT) methods. Functional magnetic resonance imaging (fMRI) is relatively unstudied in this area, though there is a growing body of literature on its use to evaluate DBS effects in other disorders including Parkinson’s disease, Essential Tremor, Tourette’s syndrome, obsessive-compulsive disorder, epilepsy, and pain (see fMRI review by Loh et al. 2022) [3]. This review will survey the existing literature in functional imaging of the effects of DBS in dystonia (Table 1) and highlight potential directions for future studies.
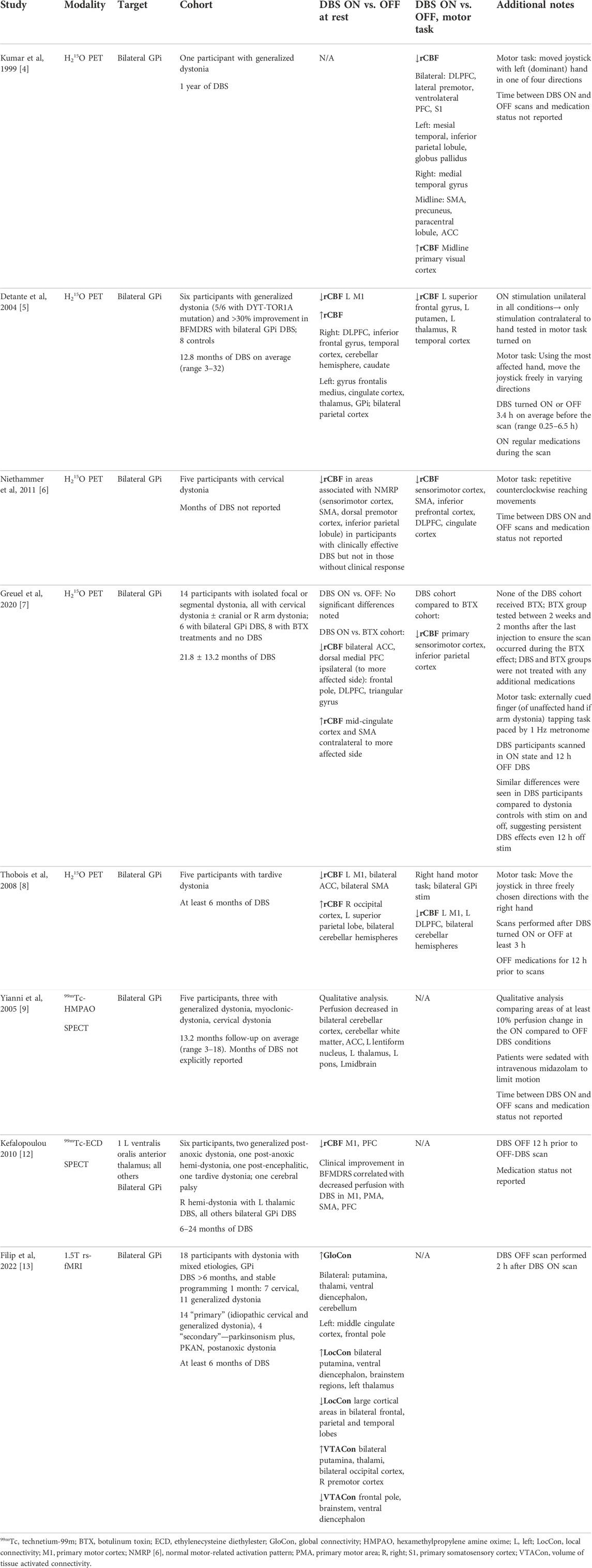
TABLE 1. Functional imaging studies of deep brain stimulation in dystonia.
PET studies
The earliest PET study of DBS in dystonia [4] evaluated one participant with idiopathic generalized dystonia who had undergone bilateral GPi DBS 1 year prior with 65% improvement in the Burke Fahn Marsden Dystonia Rating Scale (BFMDRS). H215O PET scans were completed with DBS in clinical settings, at half-amplitude, and fully off. Areas of activation were evaluated while performing a motor task requiring the manipulation of a joystick in one of four directions as compared to a rest state. This comparison was repeated in both DBS-on and DBS-off conditions. Compared to the DBS-off condition, DBS-on at half amplitude was associated with relatively reduced regional cerebral blood flow (rCBF) in widespread areas, particularly frontal cortical regions including the dorsolateral prefrontal cortex (DLPFC), lateral premotor, ventrolateral PFC, and primary sensory areas. Additionally noted were reduced rCBF in the left globus pallidus, supplementary motor area (SMA), precuneus, and anterior cingulate cortex (ACC). The only area of increased activation was in the midline primary visual cortex. The authors note a similar activation pattern with DBS at full clinical stimulation settings but with reduced statistical significance which they surmised was related to a significant head movement artifact during that scan.
Detante et al subsequently evaluated H215O PET in a cohort of six participants with generalized dystonia (five with DYT-TOR1A-related dystonia) who had undergone bilateral GPi DBS and eight healthy controls [5]. During rest and task-based study evaluations, they kept GPi stimulation on only in the hemisphere contralateral to the hand most affected by dystonia. In the rest condition, DBS-on compared to DBS-off (average 3.4 h washout) was associated with reduced rCBF in the left primary motor cortex but increased rCBF in right DLPFC, inferior frontal gyrus, temporal cortex, right cerebellar hemisphere, and right caudate. During a motor task in which the most affected hand was used to move a joystick freely in varying directions, DBS-on compared to DBS-off comparisons revealed group-level reduction of rCBF in the left superior frontal and right temporal gyri. However, these latter effects should be carefully interpreted as brain images were flipped along the x-axis for three left-hand use participants to mimic right-hand use to allow for pooling of the data among all participants.
Another small study with five participants with cervical dystonia evaluated H215O PET while DBS was turned on and off [6]. In participants with good clinical response, but not in those without, active stimulation in a non-motor sensory condition was associated with significantly reduced rCBF in the sensorimotor cortex, SMA, dorsal PMC, inferior parietal lobule, and bilateral cerebellar vermis and cerebellar hemispheric regions. During a motor task involving repetitive counterclockwise reaching movements, active stimulation resulted in reduced rCBF in the sensorimotor cortex, SMA, inferior PFC, DLPFC, and cingulate cortex. These findings highlight the importance of contextualizing imaging findings with clinical outcomes to disentangle functional changes that are associated with desired DBS effects from those that are not.
In a more recent study, Greuel and colleagues evaluated 14 participants with isolated focal or segmental dystonia (all with cervical dystonia, and some also with concurrent cranial or brachial dystonia) [7]. Two cohorts were studied, one with six participants who had undergone bilateral GPi DBS 21.8 ± 13.2 months prior, and one with eight participants who received ongoing botulinum toxin (BTX) injections without DBS. Within the DBS cohort, no significant differences in a rest condition were seen in a comparison of DBS-on versus DBS-off after a 12 h washout period. With stimulation on, the DBS group as compared to the BTX group showed reduced rCBF in the frontal-predominant cortical regions, including bilateral dorsal medial PFC and ACC, in addition to the DLPFC, triangular gyrus, and frontal pole ipsilateral to the more affected side. These same differences were seen between the cohorts even with DBS off. In contrast, there were several regions in which rCBF differed in the DBS-on condition compared to the off condition. Precuneus activity was reduced in the DBS-on, but not the DBS-off condition as compared to the BTX cohort. Conversely, increased rCBF was seen in the mid-cingulate cortex and SMA contralateral to the more affected side with DBS on but not off as compared to the BTX cohort. During a finger-tapping task, the DBS cohort showed reduced rCBF in the primary sensorimotor and inferior parietal cortices in both the DBS-on and off conditions as compared to the injection cohort.
Of particular interest in this study was the finding of prefrontal network differences in the DBS cohort compared to the BTX cohort even after a 12 h washout of DBS. One could interpret this to indicate that, in contrast to the BTX cohort, the DBS cohort experienced enduring network changes related to chronic stimulation that did not revert to a baseline pattern in only 12 h. An alternative interpretation might be that these differences were mediated by response to BTX treatment. Comparison to a control group that had received neither intervention might clarify these interpretations. A longitudinal study design in which participants had baseline evaluations prior to DBS implantation or BTX injection would offer further clarity.
Another H215O PET study in a cohort of five participants with tardive dystonia who had undergone bilateral GPi DBS demonstrated increased rCBF in the right occipital cortex, left superior parietal lobe, and bilateral cerebellar hemispheres when DBS was turned on versus off (3 h washout) in a rest condition [8]. In the same rest condition, areas of reduced rCBF were noted in the left primary motor cortex, bilateral ACC, and SMA. In a motor task using the right hand to move a joystick, reduced rCBF was noted in the left primary motor cortex, left DLPFC, and bilateral cerebellar hemispheres.
SPECT studies
There are two published studies of DBS in dystonia using technetium-99m radioisotopes. Yianni and colleagues scanned five participants with variable phenotypes of dystonia who had undergone bilateral GPi DBS, including three with generalized dystonia, one with myoclonus-dystonia, and one with cervical dystonia [9]. All were DYT-TOR1A negative, with no additional genetic testing reported. In a qualitative evaluation, they noted areas of at least 10% change in perfusion when comparing scans administered in a DBS-on versus off state. DBS-off states were not defined by a common amount of time for DBS washout, but instead as the “worst clinical state” with stimulation inactivated. The amount of time elapsed since DBS surgery was not specified. Participants were sedated with midazolam for all image acquisitions to minimize dystonic movements. Results showed areas of reduced perfusion in the DBS-on compared to DBS-off conditions in the bilateral ACC and cerebellar cortex and white matter, and the left lentiform nucleus, thalamus, pons, and midbrain. At a 15% threshold, the same areas showed reduced perfusion except the brainstem and right cerebellar regions. Although present in both scan conditions, sedation with midazolam could be confounding these findings given its prior association with reduced rCBF in the PFC [10] as well as dose-dependent rCBF reductions in the insula, cingulate gyrus, thalamus, and parietal and temporal regions [11].
Kefalopoulou et al. scanned six participants with varying forms of acquired dystonia, including three with post-anoxic dystonia (two generalized, one right hemi-dystonia), one with post-encephalitic dystonia, one with tardive dystonia, and one with cerebral palsy [12]. All had undergone bilateral GPi DBS with the exception of one participant with right hemi-dystonia who was implanted in the left ventralis oralis anterior thalamus. Scans were acquired between 6 and 24 months post-operatively. Imaging in a rest condition with DBS turned on as compared to DBS turned off after a 12 h washout period demonstrated reduced rCBF in the primary motor cortex and PFC. It was noted that there was clinical variability in response to DBS, with two participants experiencing excellent DBS response, two with moderate but present response, and two without clear clinical benefit. Clinical improvement as measured by BFMDRS was significantly correlated with decreased perfusion in the primary motor, premotor, supplementary motor, and prefrontal cortices.
fMRI studies
Only one fMRI study has been published in the context of DBS for dystonia [13]. Filip and colleagues evaluated 18 participants with dystonia from mixed etiologies. Eleven had generalized dystonia, seven had cervical dystonia, and four acquired dystonia (one in the context of Pantothenate kinase-associated neurodegeneration [PKAN], one with post-anoxic dystonia, and two in the context of parkinsonism). All the participants had undergone bilateral GPi DBS which had been activated at least 6 months prior to study inclusion and had stable programming parameters. Resting-state fMRI was performed with stimulation turned on versus off (scanned approximately 2 h apart) and two connectivity measures were evaluated including 1) Global Connectivity (GloCon), a measure of voxel/vertex-wise weighted degree centrality [14] that describes how connected and central individual brain regions are across the whole brain, and 2) Local Connectivity (LocCon), a repeated measure of Kendall’s coefficient concordance [15] over cortical and subcortical defined regions of 27 neighboring voxels that describes how synchronous the local voxel-wise fMRI timeseries are. A general linear model applied to the parcellated GloCon and LocCon brain maps to contrast DBS-on and DBS-off conditions revealed that DBS-on was associated with decreased LocCon in many cortical regions, including the bilateral frontal, parietal and temporal cortices. Increased LocCon was seen in the bilateral putamina, ventral diencephalon, brainstem regions, and the left thalamus. Across the whole-brain, increased GloCon was noted in the bilateral putamina, thalami, ventral diencephalon and cerebellum, left middle cingulate cortex, and left frontal pole, indicating that these areas became more connected and centralized to the entire brain network when DBS was turned on. When compared to healthy control data, the study authors noted that patients’ GloCon patterns in the DBS-on state were more similar to that of the healthy controls than the DBS-off state, with increased GloCon in the putamen, ventral diencephalon, cerebellum, and several cortical regions. In contrast, DBS appeared to alter LocCon in a direction opposite to the pattern seen in controls in the bilateral putamina and several cortical regions, although this effect was smallest for the best responders. Clinical improvement was otherwise found to correlate with increased GloCon over large cortical regions, the bilateral putamina, ventral diencephalon, cerebellum, and brainstem. Taken together, this work underscores the value in measuring functional response to DBS at multiple scales. The discordance between LocCon and GloCon with respect to normalization of the networks is an informative finding that, as the authors point out, could reflect variations in outcome due to pathologic differences. Alternatively, with further optimization of DBS settings, it is possible that both measures might move more toward those of healthy controls. Further insights can be gained through more densely sampled clinical assessments to monitor the rate of change in symptoms up to the time of imaging, or by mapping the functional response to multiple DBS settings and contrasting network normalization effects.
Discussion and future directions
While the studies represented in this review generally have very small sample sizes, several important patterns are worth highlighting. First, there are often different changes in functional activation when evaluated at rest as compared to motor task conditions. Such a divergence may be expected given the differences between these conditions, with one experimentally inducing performance-related motor activation, and the other in the absence of any intended movement. Second, several general patterns emerge in the evaluation of these studies collectively. The majority show reduced functional activation patterns with DBS turned on compared to off during motor tasks involving the hands, particularly in the frontal cortical regions including the PFC and sensorimotor cortex, though some studies showed reduced activation in the temporal, parietal, ACC, and cerebellar regions, as well as the thalamus, putamen, and globus pallidum. Only one study showed any region of increased rCBF during a motor task (Kumar et al with midline primary visual cortex in one participant only [4]). This may reflect changes in a common final pathway with GPi stimulation (which was overwhelmingly the target represented in these studies with the exception of a single participant with thalamic stimulation [12]).
In the surveyed studies, DBS effects during rest conditions were variable, though many did show reduced activation in the primary motor cortex and other frontal cortical areas. Other studies showed that when turned on, DBS was associated with increased rCBF in other frontal cortical areas, the cerebellum, mid-cingulate cortex, and SMA. Only one study (Detante et al. [5]) found increased rCBF in subcortical structures at rest with DBS on, including the thalamus and GPi. Of note, several studies found that only those participants who experienced clinical benefit from DBS showed decreased perfusion during a rest condition with stimulation turned on.
Differences between studies might be mediated by many factors including heterogeneous cohorts (including participants with focal or segmental isolated dystonia, genetically characterized and idiopathic generalized dystonia, acquired dystonia including post-anoxic injury, cerebral palsy, or dystonia in the context of parkinsonism), differing study conditions (e.g., sedation with midazolam or no sedation), differing washout periods for the DBS-off condition, differences in length of time elapsed since DBS implantation, differences in targeting accuracy, different rates of clinical benefit among the cohorts, and different imaging techniques. Small sample sizes also add caution to the interpretation and generalizability of study findings.
It is difficult to compare the only fMRI study in the review with the others, particularly given its use of connectivity evaluations. The scarcity of such studies reflects the safety concerns that have existed historically regarding MRI in individuals with DBS hardware. Such concerns relate to three documented cases of MRI-related injury that were partially responsible for the 2005 Food and Drug Administration warning regarding MRI in patients with DBS devices [16]. Only in 2016 did Medtronic (at that time the only manufacturer with FDA approval of DBS hardware in the US) receive FDA approval for conditional full-body MRI, at which time the acquisition of fMRI with active DBS became a technical possibility. However, specific limits were specified, such as the total duration of scanning time, use of limited MRI parameters, and permissibility of only certain active DBS parameters (including bipolar stimulation settings only). Prior to 2016, there were only a handful of studies of active DBS in Parkinson’s disease, Essential Tremor, pain, and Obsessive Compulsive Disorder, with an increasing number subsequent to 2016 [3].
To date, the study by Filip and colleagues [13] represents the only published fMRI study of active DBS in dystonia. Hopefully, there will be additional fMRI studies of DBS in dystonia in the future to evaluate the replicability of the network rearrangement patterns seen in the study. The heterogeneity of the study is a potential confound, particularly the inclusion of acquired and isolated forms of dystonia in the same cohort, as baseline connectivity patterns and response to DBS may vary significantly among them.
Several potential confounds must be addressed when considering the design of future functional neuroimaging studies in the mechanisms of DBS in dystonia. First, if dystonic movements are present at the time of imaging, resulting changes in functional activation patterns may be related to the movements themselves rather than specific to changes associated with the dystonia (i.e., the same pattern might be seen with a similar voluntary movement, as opposed to dystonic movements). Relative to this concern, evaluations at rest may therefore have cleaner interpretations. Nonetheless, in some cases, such confounds may be addressed by a control motor task if possible. On an intra-subject basis, this might involve mimicking the same movement with an unaffected contralateral side. Alternatively, an inter-subject comparison might be performed with a healthy control. An additional confound may occur if dystonic movements affect the head or neck in the midst of scanning, thereby introducing motion artifacts that may reduce image quality and degrade the interpretability and fidelity of both structural and functional imaging analysis. Targeted inclusion and exclusion criteria might be considered to circumvent this concern, such as excluding cervical dystonia participants who also have overlying head tremors and including only those with a pure posturing phenotype. However, this approach runs the risk of creating selection biases which may skew results, particularly if there are intrinsic differences in underlying physiology in cervical dystonia with and without tremors as demonstrated in some settings [17]. A better approach may be evaluating individual potential cervical dystonia participants with and without overlying head tremors and the degree of head movement they exhibit while lying supine, excluding only those with frequent and severe enough movements in this position to preclude adequate scan quality. Given the very small sample sizes, one might even consider using MR-compatible wearables to measure tremors during fMRI acquisition followed by post hoc regression-based denoising methods. While sedation can prevent motion-related confounds altogether, anesthesia is also associated with independent confounding effects. Despite these inevitable tradeoffs and challenges that come with executing functional imaging studies of DBS modulation, efforts must be made to make conditions otherwise as identical as possible to attempt to isolate functional changes specifically related to stimulation and not other variables.
Important insights into the mechanisms of DBS benefit in dystonia might be gleaned particularly from well-designed longitudinal studies which include both baseline pre-operative scans and post-operative scans at defined intervals with DBS on and off. It would be important to include clinical measures to correlate the DBS effect with imaging changes. Appropriate outcome measures should be considered for the study cohort to ensure that the clinical scales employed are adequate to capture changes in the body region most affected by dystonia. This is important as scales that evaluate multiple body regions, such as the Burke Fahn Marsden Dystonia Rating Scale [18], may not adequately reflect the degree of clinical change in one specific body region in a cohort of focal dystonia, as even large focal changes may be washed out in the overall score. Study evaluation parameters should be standardized as much as possible, including length of time elapsed since implantation, amount of time for DBS washout for DBS-off condition scans, and more homogeneous cohorts. Given the rarity of dystonia and far fewer people annually receiving DBS for dystonia than for other conditions such as Essential Tremor or Parkinson’s disease, there will always be challenges in recruiting larger cohorts for these studies. Multi-center collaborative studies are one important approach to overcome this limitation. Longitudinal study designs are another, given the considerable increase in statistical power and insight with repeated observations over time even at the intra-participant level.
A growing body of literature has explored associations between connectivity patterns and DBS outcomes using the volume of tissue activated (VTA) by stimulation as seed regions to probe functional and structural connectivity patterns within normative atlases [19, 20]. These approaches have undoubtedly advanced the general understanding of regional connectivity in DBS. However, studies evaluating actual connectivity patterns of dystonia patients with DBS offer important additional information, given the possibility that connectivity may deviate from that of healthy controls as part of the dystonia endophenotype and as a result of network rearrangement from DBS. With the increasing use of fMRI with DBS, this will likely represent an important area of study in the future.
Author contributions
IB—article design, writing initial draft, reviewing and revising. MM—article design, reviewing and revising. All authors contributed to the article and approved the submitted version.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Jimenez-Shahed, J. Device profile of the percept PC deep brain stimulation system for the treatment of Parkinson’s disease and related disorders. Expert Rev Med Devices (2021) 18:319–32. doi:10.1080/17434440.2021.1909471
2. Schirinzi, T, Sciamanna, G, Mercuri, NB, and Pisani, A. Dystonia as a network disorder: a concept in evolution. Curr Opin Neurol (2018) 31:498–503. doi:10.1097/WCO.0000000000000580
3. Loh, A, Gwun, D, Chow, CT, Boutet, A, Tasserie, J, Germann, J, et al. Probing responses to deep brain stimulation with functional magnetic resonance imaging. Brain Stimulat (2022) 15:683–94. doi:10.1016/j.brs.2022.03.009
4. Kumar, R, Dagher, A, Hutchison, WD, Lang, AE, and Lozano, AM. Globus pallidus deep brain stimulation for generalized dystonia: clinical and PET investigation. Neurology (1999) 53:871–4. doi:10.1212/wnl.53.4.871
5. Detante, O, Vercueil, L, Thobois, S, Broussolle, E, Costes, N, Lavenne, F, et al. Globus pallidus internus stimulation in primary generalized dystonia: a H215O PET study. Brain (2004) 127:1899–908. doi:10.1093/brain/awh213
6. Niethammer, M, Carbon, M, Argyelan, M, and Eidelberg, D. Hereditary dystonia as a neurodevelopmental circuit disorder: evidence from neuroimaging. Neurobiol Dis (2011) 42:202–9. doi:10.1016/j.nbd.2010.10.010
7. Greuel, A, Pauls, KAM, Koy, A, Südmeyer, M, Schnitzler, A, Timmermann, L, et al. Pallidal deep brain stimulation reduces sensorimotor cortex activation in focal/segmental dystonia. Mov Disord (2020) 35:629–39. doi:10.1002/mds.27970
8. Thobois, S, Ballanger, B, Xie-Brustolin, J, Damier, P, Durif, F, Azulay, JP, et al. Globus pallidus stimulation reduces frontal hyperactivity in tardive dystonia. J Cereb Blood Flow Metab (2008) 28:1127–38. doi:10.1038/sj.jcbfm.9600610
9. Yianni, J, Bradley, K, Soper, N, O'Sullivan, V, Nandi, D, Gregory, R, et al. Effect of GPi DBS on functional imaging of the brain in dystonia. J Clin Neurosci (2005) 12:137–41. doi:10.1016/j.jocn.2004.05.010
10. Reinsel, RA, Veselis, RA, Dnistrian, AM, Feshchenko, VA, Beattie, BJ, and Duff, MR. Midazolam decreases cerebral blood flow in the left prefrontal cortex in a dose-dependent fashion. Int J Neuropsychopharmacol (2000) 3:117–27. doi:10.1017/S1461145700001814
11. Veselis, RA, Reinsel, RA, Beattie, BJ, Mawlawi, OR, Feshchenko, VA, DiResta, GR, et al. Midazolam changes cerebral blood flow in discrete brain regions: an H2(15)O positron emission tomography study. Anesthesiology (1997) 87:1106–17. doi:10.1097/00000542-199711000-00015
12. Kefalopoulou, Z, Paschali, A, Markaki, E, Ellul, J, Chroni, E, Vassilakos, P, et al. Regional cerebral blood flow changes induced by deep brain stimulation in secondary dystonia. Acta Neurochir. (Wien) (2010) 152:1007–14. doi:10.1007/s00701-010-0612-y
13. Filip, P, Jech, R, Fečíková, A, Havránková, P, Růžička, F, Mueller, K, et al. Restoration of functional network state towards more physiological condition as the correlate of clinical effects of pallidal deep brain stimulation in dystonia. Brain Stimulat (2022) 15:1269–78. doi:10.1016/j.brs.2022.08.025
14. Craddock, RC, and Clark, DJ. Optimized implementations of voxel-wise degree centrality and local functional connectivity density mapping in AFNI. GigaScience (2016) 5. doi:10.1186/s13742-016-0147-0-d
15. Zang, Y, Jiang, T, Lu, Y, He, Y, and Tian, L. Regional homogeneity approach to fMRI data analysis. NeuroImage (2004) 22:394–400. doi:10.1016/j.neuroimage.2003.12.030
16. Boutet, A, Chow, CT, Narang, K, Elias, GJB, Neudorfer, C, Germann, J, et al. Improving safety of MRI in patients with deep brain stimulation devices. Radiology (2020) 296:250–62. doi:10.1148/radiol.2020192291
17. Sedov, A, Usova, S, Semenova, U, Gamaleya, A, Tomskiy, A, Beylergil, SB, et al. Pallidal activity in cervical dystonia with and without head tremor. Cerebellum Lond Engl (2020) 19:409–18. doi:10.1007/s12311-020-01119-5
18. Burke, RE, Fahn, S, Marsden, CD, Bressman, SB, Moskowitz, C, and Friedman, J. Validity and reliability of a rating scale for the primary torsion dystonias. Neurology (1985) 35:73–7. doi:10.1212/wnl.35.1.73
19. Reich, MM, Horn, A, Lange, F, Roothans, J, Paschen, S, Runge, J, et al. Probabilistic mapping of the antidystonic effect of pallidal neurostimulation: a multicentre imaging study. Brain (2019) 142:1386–98. doi:10.1093/brain/awz046
20. Okromelidze, L, Tsuboi, T, Eisinger, RS, Burns, MR, Charbel, M, Rana, M, et al. Functional and structural connectivity patterns associated with clinical outcomes in deep brain stimulation of the globus pallidus internus for generalized dystonia. Am J Neuroradiol (2020) 41:508–14. doi:10.3174/ajnr.A6429
Keywords: dystonia, deep brain stimulation, functional imaging, fMRI, PET
Citation: Bledsoe IO and Morrison MA (2023) Functional imaging of deep brain stimulation in dystonia: a review. Dystonia 2:11440. doi: 10.3389/dyst.2023.11440
Received: 03 April 2023; Accepted: 14 September 2023;
Published: 06 October 2023.
Edited by:
Aasef Shaikh, Case Western Reserve University, United StatesCopyright © 2023 Bledsoe and Morrison. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ian O. Bledsoe, aWFuLmJsZWRzb2VAdWNzZi5lZHU=